GOLDEN RICE IS A PART OF SOLUTION
Biofortified rice as a contribution to the alleviation of life-threatening micronutrient deficiencies in developing countries.

The second best approach, especially for those who cannot afford a varied diet, is by way of nutrient-dense staple crops. Sweet potatoes, for example, are available as varieties that are either rich or poor in provitamin A. Those producing and accumulating provitamin A (orange-fleshed sweet potatoes) are called biofortified*, as opposed to the white-fleshed sweet potatoes, which do not accumulate provitamin A. In this case, what needs to be done is to introduce the biofortified varieties to people used to the white-fleshed varieties. Unfortunately, there are no natural provitamin A-containing rice varieties.
Rice plants produce β-carotene (provitamin A) in green tissues but not in the endosperm (the edible part of the seed). The outer coat of the dehusked grains—the so-called aleurone layer—contains a number of valuable nutrients, e.g. vitamin B and nutritious fats, but no provitamin A. These nutrients are lost with the bran fraction in the process of milling and polishing. While it would be desirable to keep those nutrients, the fatty component is affected by oxidative processes that make the grain turn rancid. Thus, unprocessed rice—also known as brown rice—is not apt for long-term storage.
Even though all required genes to produce provitamin A are present in the grain, some of them are turned off during development. In rice-based societies, the absence of β-carotene in rice grains manifests itself in a marked incidence of blindness, disease susceptibility and premature death of small children.
Rice plants produce β-carotene (provitamin A) in green tissues but not in the endosperm (the edible part of the seed). The outer coat of the dehusked grains—the so-called aleurone layer—contains a number of valuable nutrients, e.g. vitamin B and nutritious fats, but no provitamin A. These nutrients are lost with the bran fraction in the process of milling and polishing. While it would be desirable to keep those nutrients, the fatty component is affected by oxidative processes that make the grain turn rancid. Thus, unprocessed rice—also known as brown rice—is not apt for long-term storage.
Even though all required genes to produce provitamin A are present in the grain, some of them are turned off during development. In rice-based societies, the absence of β-carotene in rice grains manifests itself in a marked incidence of blindness, disease susceptibility and premature death of small children.
SCIENCE BEHIND THE GOLDEN RICE
Filling in the gaps
Golden Rice technology is based on a simple principle. Notwithstanding the fact that rice plants synthesise β-carotene in vegetative tissues but not in the grain, all but two steps of the biosynthetic pathway are present in the grain. By addition of only two genes, phytoene synthase (psy) and phytoene desaturase (crt I), the pathway is reconstituted and β-carotene is consequently accumulated in the endosperm, ie the edible part of the grain.
 Carotenoids and their derivatives include a vast number of molecules and accordingly a great number of enzymes and cofactors. Compounds derived from this important pathway include plant hormones, like abcisic acid and gibberellins, vitamin E, part of the chlorophyll molecule, and many natural pigments. A small number of carotenoids have provitamin A activity. |
The underlying science in more detail
All plant tissues that accumulate high levels of carotenoid have a mechanism for carotenoid sequestration including crystallisation, oil deposition, membrane proliferation or protein-lipid sequestration. The non-carotenogenic starchy rice endosperm is very low in lipid and apparently lacks any such means for carotenoid deposition. Another restriction in Golden Rice could have been precursor supply. Also, many people believed that the whole carotenoid biosynthetic pathway—composed of many steps—was completely absent in the endosperm.For these reasons the generation of Golden Rice went through a long lag phase, until enough scientific data had been accumulated, which encouraged Peter Beyer and Ingo Potrykus to work together and try to engage in this seeminlgy unattainable endeavour. Their breakthrough showed that only two transgenes were required to turn Golden Rice into a reality, and that the feared restrictions described above did not apply (Ye et al., 2000). The first transgene encodes phytoene synthase (PSY), which utilises the endogenously synthesised geranylgeranyl-diphosphate to form phytoene, a colorless carotene with a triene chromophore (Burkhardt et al., 1997). The second encodes a bacterial carotene desaturase (CRTI) that introduces conjugation by adding four double bonds. The combined activity of PSY and CRTI leads to the formation of lycopene, which is a red compound due to its undecaene chromophore. Lycopene has never been observed in any rice transformant and different genetic backgrounds. Instead, alpha- and beta-carotene are found together with variable amounts of oxygenated carotenoids such as lutein and zeaxanthin. The carotenoid pattern observed in the endosperm revealed that the pathway proceeded beyond the end point expected from the enzymatic action of the two transgenes alone. A detail analysis of the underlying mechanism was recently published by co-workers from Peter Beyer's lab in the journal Plant Physiology (Schaub et al., 2005). Their findings are explained in some detail below.
 The precursor molecule for carotenoid biosynthesis is geranylgeranyl diphosphate (GGDP). Horizontal bars delimit the steps of the carotenoid biosynthetic pathway that were overcome using the two transgenes phytoene synthase (PSY) and the multifunctional bacterial carotene desaturase (CRTI), rather than the two plant desaturases PDS and ZDS. |
One explanation is that enzymes downstream along the pathway, such as lycopene cyclases (LCYs) and alpha- and beta-carotene hydroxylases (HYDs) are are being produced in wild-type rice endosperm, while PSY and one or both of the plant carotene desaturases—phytoene desaturase (PDS) and ζ-carotene desaturase (ZDS)—are not. Synthesis of lycopene by PSY and CRTI in transgenic plants provides the substrate for these downstream enzymes and consequently enables the formation of observed products. The fact that a PSY transgene alone led to phytoene accumulation but not to desaturated products (Burkhardt et al., 1997) is evidence for the absence of at least one active desaturase, namely PDS. Similarly, the expression of CRTI alone did not produce any color in rice endosperm, because of the lack of PSY activity.The alternative explanation involves feedback induction of endogenous carotenoid biosynthetic genes as a result of the presence of the transgenes. This was shown to be the case in tomato (Lycopersicon esculentum) fruit upon expression of CRTI alone. Again, beta-carotene rather than lycopene increased, and endogenous carotenoid biosynthetic genes were shown to be upregulated, except for PSY, which was repressed (Römer et al., 2000). Such CRTI-dependent up-regulation may be based on the fact that the plant desaturases, PDS and ZDS, together produce a tetra-cis configured form of lycopene, termed prolycopene (Bartley et al., 1999), which is subsequently isomerized to the trans form by a recently identified isomerase, lycopene-cis- transisomerase (CRTISO; Isaacson et al., 2002, 2004; Park et al., 2002). In contrast, the bacterial CRTI leads to the exclusive formation of the all-trans form of lycopene. Likewise, an increased formation of specific carotenogenic mRNAs and proteins was consistently observed in daffodil flowers upon artificial accumulation of all-trans lycopene after inhibition of LCY, and the total carotenoid content was elevated (Al-Babili et al., 1999).
Plants containing a constitutively expressed crtI transgene were used to explain the Golden Rice phenotype. This was achieved by determining changes in the expression levels of endogenous carotenoid biosynthetic genes and their products. It was conceivable that the presence of CRTI provoked transcriptional activation of the carotenoid biosynthetic genes in the target tissue by a feedback regulatory loop. The loop could be initiated by the absence of tetra-cis-lycopene, normally produced by the two plant desaturases PDS and ZDS, or the constitutive presence of all-trans-lycopene, mediated by CRTI. Lycopene isomers would not only represent orthodox intermediates but would also exert a regulatory role. The newly identified carotenoid pathway enzyme of plants and cyanobacteria, CRTISO (Isaacson et al., 2002; Park et al., 2002), could have played an additional role as regulator. In such a scenario, Golden Rice would have been yellow due to the transcriptional activation of the entire carotenoid biosynthetic pathway.
In an alternative scenario, active downstream enzymes, such as cyclases and hydroxylases, might be present in wild-type rice endosperm, thus explaining the processing of lycopene immediately after synthesis by the introduced upstream transgenes. Wild- type rice endosperm displays low levels of all mRNAs required for xanthophyll formation, ie PSY, PDS, ZDS, CRTISO, β-LCY, ε;-LCY, β-HYD, and ε-HYD. Among these and given the sensitivity of the PCR method used, the PSY transcript was effectively absent and this is consistent with the fact that introduction of PSY is required—but not sufficient—to produce the golden phenotype. The expression of PSY alone results in the accumulation of phytoene (Burkhardt et al., 1997).
The essential requirement for CRTI apparently conflicts with the presence of PDS and ZDS transcripts in wild-type endosperm, as shown by TaqMan real-time PCR. This could be due to low level protein expression and enzymatic activity rather than mRNA levels. Due to the low expression, the complicated underlying reaction mechanisms of PDS and ZDS and the unavailability of radioactive carotene substrates, the investigations were done using a transgenic approach rather than in vitro reactions. The endosperm- specific expression of the PDS/ZDS system—instead of CRTI—in rice endosperm resulted in the formation of comparable levels of colored carotenoids. Thus, the rice endosperm provides the complex requirements for the activity of the plant desaturases.
PDS requires a redox chain, employing quinones, a quinone-reductase, and molecular oxygen as a terminal electron acceptor (Beyer et al., 1989; Mayer et al., 1990; Nievelstein et al., 1995) to which it is linked via an oxidase identified through the immutans mutation of Arabidopsis (for review, see Kuntz, 2004). This redox pathway is especially important in non- green carotenoid-bearing tissues like endosperm, while the photosynthetic electron transport is thought to play an analogous role in chloroplasts. Therefore, one interpretation is that colored carotenoids do not form in the endosperm with PSY as the only transgene because the expression of the rice PDS/ZDS system is too low. It has been shown that, in contrast to PSY, CRTI is not rate limiting and is capable of desaturating large amounts of phytoene, thereby increasing beta-carotene accumulation (Paine et al., 2005).
The primary sequence of CRTI is unrelated to the plant-type desaturases. This might explain simpler co-factor requirements and may therefore be more effective in rice endosperm than the plant-type desaturases. On the other hand, CRTISO, which is also active in rice endosperm seems to have originated from CRTI in evolution (Isaacson et al., 2002; Park et al., 2002).
PSY, PDS, and ZDS expression in rice endosperm installed the poly-cis pathway of carotene desaturation, but the beta-carotene formed was predominantly in the all-trans form, accompanied by the typical pattern of cis-isomers, the 9-cis form being the most abundant. Furthermore, endosperm from transgenic plants relying on CRTI-mediated desaturation yielded the identical isomer ratio of beta-carotenes, indicating that rice CRTISO catalyses the formation of these isomers at this specific ratio, consistent with its in vitro activity (Isaacson et al., 2004). The activity of CRTISO is also crucial with respect to the formation of cyclic end groups. In its absence, cyclic carotenoids and derived xanthophylls would not form in non-green tissues, as has been shown with the tangerine mutation in tomato fruit and in etioplasts from the Arabidopsisccr2 mutant, both lacking functional CRTISO (Isaacson et al., 2002; Park et al., 2002).
Clearly, LCY activities and the activities of the divergent class of ß-HYD (for review, see Tian and DellaPenna, 2004) rely on the expression of the respective rice genes in the endosperm. In all rice genetic backgrounds tested so far, complementation with these activities is not required to proceed down the pathway. Moreover, the activity of rice LCYs never proved to be rate limiting, since lycopene did not accumulate. Thus, Golden Rice is yellow because of the activity of intrinsic rice cyclases. TaqMan real-time PCR analyses with RNA isolated from transgenic rice endosperm did not give any indication of a feedback regulatory loop that may affect the expression of rice carotenoid biosynthetic genes, because all the corresponding mRNA levels remained unchanged when compared to the wild-type.
The situation was similar in leaves of rice and Arabidopsis expressing CRTI constitutively, despite an increase in xanthophylls. In both cases, lutein decreased, partially compensated by an increase in ß-carotene and its derived xanthophylls. This was also the case in tobacco BY2 cells transformed with constitutively expressed CRTI, but here no comparable change in carotenoid composition was found, probably because BY2 cells do not accumulate lutein and zeaxanthin. The change in the xanthophyll ratio observed in rice and Arabidopsis leaves varied in an event-dependent manner and was inversely correlated to the CRTI expression level. The flux of substrate into either branch of xanthophyll formation is controlled by the two LCYs, which convert lycopene into β- carotene (β,β-carotene) and α-carotene (β ε-carotene). However, no significant changes in the levels of the trancripts of the cyclases were found that would mirror the change in carotenoid composition. It must therefore be assumed that the state of geometric isomerism in lycopene has a certain impact on the probability of β- or ε;-ring formation.
The lack of CRTISO in mutants of tomato and Arabidopsis also led to a predominance of the beta-carotene-derived xanthophylls in leaves (Isaacson et al., 2002; Park et al., 2002). Thus, the observed effect is most likely at the level of enzymatic catalysis and geometric isomerism of lycopene, but not at the level of gene expression. This effect is evident only in tissues exhibiting carotenoid biosynthesis in the wild-type, such as in leaves. It cannot occur in transformed endosperm, which lacks carotenoids in the wild- type.
The observed decrease in lutein in leaves might, in theory, have a negative impact on photosynthetic performance. Lutein, the most abundant xanthophyll, is primarily attached to the light-harvesting complex II. Using mutants of Arabidopsis, it has been shown that a decrease in lutein content leads to a reduction of PSII antenna size (Lokstein et al., 2002). This suggests that lutein has the ability to optimize antenna structure and stability to ensure efficient light harvesting. Therefore, we chose to express crtI under the control of a tissue-specific promoter in newer versions of Golden Rice (Paine et al., 2005; Al-Babili et al., submitted; TTC Hoa and P Schaub, unpublished results).
These efforts led to the development of what we could call the first generation of Golden Rice (after the proof of concept), also known as SGR1. This version only contained the phytoene synthase (psy) gene from daffodil and the carotene desaturase (crtI) gene from the bacteriumErwinia uredovora. Further, in this version both genes were expressed only in the rice endosperm. The levels of carotenoids obtained in the greenhouse were not very unlike the precursor version (1-2 µg/g) but in the field production and accumulation amounted to an average of 6 µg/g. This might have been due to improved growth conditions and to the selection process that the plants underwent. This level of carotenoids contents is expected to be able to cover the recommended daily intake values for children when taking into consideration a modest intake of vegetables and fish or other animal sources (as they are doing at present).
Plants containing a constitutively expressed crtI transgene were used to explain the Golden Rice phenotype. This was achieved by determining changes in the expression levels of endogenous carotenoid biosynthetic genes and their products. It was conceivable that the presence of CRTI provoked transcriptional activation of the carotenoid biosynthetic genes in the target tissue by a feedback regulatory loop. The loop could be initiated by the absence of tetra-cis-lycopene, normally produced by the two plant desaturases PDS and ZDS, or the constitutive presence of all-trans-lycopene, mediated by CRTI. Lycopene isomers would not only represent orthodox intermediates but would also exert a regulatory role. The newly identified carotenoid pathway enzyme of plants and cyanobacteria, CRTISO (Isaacson et al., 2002; Park et al., 2002), could have played an additional role as regulator. In such a scenario, Golden Rice would have been yellow due to the transcriptional activation of the entire carotenoid biosynthetic pathway.
In an alternative scenario, active downstream enzymes, such as cyclases and hydroxylases, might be present in wild-type rice endosperm, thus explaining the processing of lycopene immediately after synthesis by the introduced upstream transgenes. Wild- type rice endosperm displays low levels of all mRNAs required for xanthophyll formation, ie PSY, PDS, ZDS, CRTISO, β-LCY, ε;-LCY, β-HYD, and ε-HYD. Among these and given the sensitivity of the PCR method used, the PSY transcript was effectively absent and this is consistent with the fact that introduction of PSY is required—but not sufficient—to produce the golden phenotype. The expression of PSY alone results in the accumulation of phytoene (Burkhardt et al., 1997).
The essential requirement for CRTI apparently conflicts with the presence of PDS and ZDS transcripts in wild-type endosperm, as shown by TaqMan real-time PCR. This could be due to low level protein expression and enzymatic activity rather than mRNA levels. Due to the low expression, the complicated underlying reaction mechanisms of PDS and ZDS and the unavailability of radioactive carotene substrates, the investigations were done using a transgenic approach rather than in vitro reactions. The endosperm- specific expression of the PDS/ZDS system—instead of CRTI—in rice endosperm resulted in the formation of comparable levels of colored carotenoids. Thus, the rice endosperm provides the complex requirements for the activity of the plant desaturases.
PDS requires a redox chain, employing quinones, a quinone-reductase, and molecular oxygen as a terminal electron acceptor (Beyer et al., 1989; Mayer et al., 1990; Nievelstein et al., 1995) to which it is linked via an oxidase identified through the immutans mutation of Arabidopsis (for review, see Kuntz, 2004). This redox pathway is especially important in non- green carotenoid-bearing tissues like endosperm, while the photosynthetic electron transport is thought to play an analogous role in chloroplasts. Therefore, one interpretation is that colored carotenoids do not form in the endosperm with PSY as the only transgene because the expression of the rice PDS/ZDS system is too low. It has been shown that, in contrast to PSY, CRTI is not rate limiting and is capable of desaturating large amounts of phytoene, thereby increasing beta-carotene accumulation (Paine et al., 2005).
The primary sequence of CRTI is unrelated to the plant-type desaturases. This might explain simpler co-factor requirements and may therefore be more effective in rice endosperm than the plant-type desaturases. On the other hand, CRTISO, which is also active in rice endosperm seems to have originated from CRTI in evolution (Isaacson et al., 2002; Park et al., 2002).
PSY, PDS, and ZDS expression in rice endosperm installed the poly-cis pathway of carotene desaturation, but the beta-carotene formed was predominantly in the all-trans form, accompanied by the typical pattern of cis-isomers, the 9-cis form being the most abundant. Furthermore, endosperm from transgenic plants relying on CRTI-mediated desaturation yielded the identical isomer ratio of beta-carotenes, indicating that rice CRTISO catalyses the formation of these isomers at this specific ratio, consistent with its in vitro activity (Isaacson et al., 2004). The activity of CRTISO is also crucial with respect to the formation of cyclic end groups. In its absence, cyclic carotenoids and derived xanthophylls would not form in non-green tissues, as has been shown with the tangerine mutation in tomato fruit and in etioplasts from the Arabidopsisccr2 mutant, both lacking functional CRTISO (Isaacson et al., 2002; Park et al., 2002).
Clearly, LCY activities and the activities of the divergent class of ß-HYD (for review, see Tian and DellaPenna, 2004) rely on the expression of the respective rice genes in the endosperm. In all rice genetic backgrounds tested so far, complementation with these activities is not required to proceed down the pathway. Moreover, the activity of rice LCYs never proved to be rate limiting, since lycopene did not accumulate. Thus, Golden Rice is yellow because of the activity of intrinsic rice cyclases. TaqMan real-time PCR analyses with RNA isolated from transgenic rice endosperm did not give any indication of a feedback regulatory loop that may affect the expression of rice carotenoid biosynthetic genes, because all the corresponding mRNA levels remained unchanged when compared to the wild-type.
The situation was similar in leaves of rice and Arabidopsis expressing CRTI constitutively, despite an increase in xanthophylls. In both cases, lutein decreased, partially compensated by an increase in ß-carotene and its derived xanthophylls. This was also the case in tobacco BY2 cells transformed with constitutively expressed CRTI, but here no comparable change in carotenoid composition was found, probably because BY2 cells do not accumulate lutein and zeaxanthin. The change in the xanthophyll ratio observed in rice and Arabidopsis leaves varied in an event-dependent manner and was inversely correlated to the CRTI expression level. The flux of substrate into either branch of xanthophyll formation is controlled by the two LCYs, which convert lycopene into β- carotene (β,β-carotene) and α-carotene (β ε-carotene). However, no significant changes in the levels of the trancripts of the cyclases were found that would mirror the change in carotenoid composition. It must therefore be assumed that the state of geometric isomerism in lycopene has a certain impact on the probability of β- or ε;-ring formation.
The lack of CRTISO in mutants of tomato and Arabidopsis also led to a predominance of the beta-carotene-derived xanthophylls in leaves (Isaacson et al., 2002; Park et al., 2002). Thus, the observed effect is most likely at the level of enzymatic catalysis and geometric isomerism of lycopene, but not at the level of gene expression. This effect is evident only in tissues exhibiting carotenoid biosynthesis in the wild-type, such as in leaves. It cannot occur in transformed endosperm, which lacks carotenoids in the wild- type.
The observed decrease in lutein in leaves might, in theory, have a negative impact on photosynthetic performance. Lutein, the most abundant xanthophyll, is primarily attached to the light-harvesting complex II. Using mutants of Arabidopsis, it has been shown that a decrease in lutein content leads to a reduction of PSII antenna size (Lokstein et al., 2002). This suggests that lutein has the ability to optimize antenna structure and stability to ensure efficient light harvesting. Therefore, we chose to express crtI under the control of a tissue-specific promoter in newer versions of Golden Rice (Paine et al., 2005; Al-Babili et al., submitted; TTC Hoa and P Schaub, unpublished results).
Golden Rice: The First Generation
The first breakthrough in the development of Golden Rice was the result of a collaboration between Peter Beyer and Ingo Potrykus and was obtained around Easter 1999 (Ye et al., Science 287:303-5, 2000). This paper provided the proof that beta-carotene could be produced in the rice grain. At the time it was still believed that, beside phytoene synthase and carotene desaturase, a third enzyme, lycopene cyclase, was needed to complement the biosynthetic pathway (see discussion and diagram above).With the proof of concept in hands, the scientists immediately proceeded to develop ways of improving the production and accumulation of carotenoids in the seed, as it was recognised that at the levels attainable at the time (1.6 µg/g) Golden Rice would not be able to fully cover the daily pro-vitamin A requirements of the target population in the absence of a more varied diet. While some population strata in SE Asia do consume more varied diets, many of the poorest do not, in fact in some rural population rice makes up more than 80% of their daily caloric intake.These efforts led to the development of what we could call the first generation of Golden Rice (after the proof of concept), also known as SGR1. This version only contained the phytoene synthase (psy) gene from daffodil and the carotene desaturase (crtI) gene from the bacteriumErwinia uredovora. Further, in this version both genes were expressed only in the rice endosperm. The levels of carotenoids obtained in the greenhouse were not very unlike the precursor version (1-2 µg/g) but in the field production and accumulation amounted to an average of 6 µg/g. This might have been due to improved growth conditions and to the selection process that the plants underwent. This level of carotenoids contents is expected to be able to cover the recommended daily intake values for children when taking into consideration a modest intake of vegetables and fish or other animal sources (as they are doing at present).
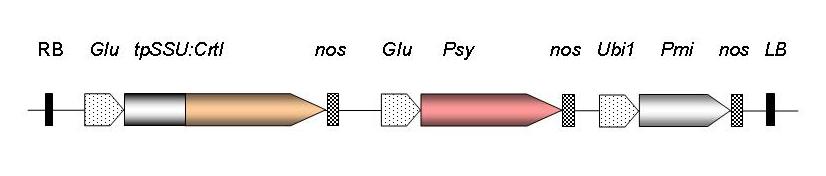 Gene construct used to generate Golden Rice. RB, T-DNA right border sequence; Glu, rice endosperm-specific glutelin promoter; tpSSU, pea ribulose bis-phosphate carboxylase small subunit transit peptide for chloroplast localisation; nos, nopaline synthase terminator; Psy, phytoene synthase gene from Narcissus pseudonarcissus (GR1) or Zea mays (GR2); Ubi1, maize polyubiquitin promoter; Pmi, phosphomannose isomerase gene from E. coli for positive selection (GR2); LB, T-DNA left border sequence. |

A new Golden Rice generation
The first generation of Golden Rice was a valuable proof of concept, but it was recognised that to combat vitamin A deficiency more efficiently higher β-carotene accumulation levels would be required. As only two transgenes are required in the process, the logical approach was to identify the bottleneck of the biosynthetic pathway and fine-tune the enzymatic activities of the two gene products involved, phytoene synthase (PSY) and carotene desaturase (CRTI). This can be done by replacing the genes with homologues from other sources or modifying their regulatory regions.In most multi-step biosynthetic pathways there is a rate-limiting step. Making a long story short, the bottleneck in this case was the enzymatic activity of PSY. After trying with PSY genes from different sources it turned out that the maize and rice genes gave the best results (Paine et al., 2005). In the process Golden Rice lines were obtained that accumulated up to 37 µg/g carotenoid of which 31 µg/g is β-carotene (as compared to the first generation Golden Rice where only 1.6 µg/g were obtained.The recommended daily allowance (RDA) of vitamin A for 1-3 year-old children is 300 µg (half the RDA is enough to maintain vitamin A at a normal, healthy level). Based on a retinol equivalency ratio for β-carotene of 12:1, half the RDA would be provided in 72 g of the new-generation Golden Rice. This is perfectly compatible with rice consumption levels in target countries, which lie at 100-200 g of rice per child per day.
Excellent post, it is very interesting to read your blog and you provide nice information. Thanks for sharing…
ReplyDeletecar transportation in Kanpur.